法政大学自然科学センター・法学部の植木紀子教授、京都産業大学生命科学部の若林憲一教授の共同研究グループは、緑藻植物門・緑藻綱・ボルボックス目に属す生物種が、急に強い光を浴びたときの繊毛運動の反応の様式が4パターンに分類できることを見出しました。そして、それぞれの種の個体の大きさと遊泳速度の解析結果から、そのパターンが、系統関係よりも、むしろ生物が生きている流体環境を示す指標である「レイノルズ数」によって分類できることを突き止めました。この結果から、繊毛運動調節機構の進化にとって、流体環境が重要な淘汰圧であることがわかりました。本研究の成果は、ヒト繊毛病の発症メカニズムの理解や、マイクロマシンの設計などに貢献することが期待されます。この成果はSpringer Nature社が発行する国際学術雑誌BMC Ecology and Evolution(2024年9月24日)に掲載されました。
本件のポイント
- 緑藻植物門・緑藻綱・ボルボックス目に属す多細胞性の藻類は、モデル単細胞緑藻として知られるクラミドモナスChlamydomonasに似た祖先単細胞生物が多細胞化して進化したと考えられている。クラミドモナスと、最大数万細胞から成る球形の多細胞緑藻ボルボックスVolvoxがよく知られるが、その間の細胞数の生物種、たとえば4細胞のテトラバエナTetrabaenaや16細胞のパンドリナPandorinaなどが現存している。そのため、化石ではなく、現存生物を用いて実験的に多細胞化進化の過程を追うことができるという、ユニークな生物群である。
- これら多細胞性の種は、クラミドモナスと同様、1細胞あたり2本の繊毛(鞭毛)が生えており、繊毛運動によって遊泳する。また、1細胞あたり1つもつ眼点と呼ばれる光受容器官で光の方向と強さを認識する。ボルボックス目藻類の多くは、光刺激に応じて繊毛の動かし方を変えて、光反応行動と呼ばれる行動を示す。
- 本研究では、ボルボックス目に属す27系統の藻類が、強い光を浴びたときに一時的に示す繊毛運動パターンの様子を観察し、「1: 波形変換」「2: 無反応」「3: 運動停止」「4: 運動方向逆転」の4つのパターンに分類できることを見出した。
- さらに、この4つのパターンは、それぞれの種の系統よりも、その大きさと遊泳速度から導かれる「レイノルズ数」に応じて決まることが示唆された。
- レイノルズ数は「慣性力」を「粘性抵抗」で割ったものである。微生物は低レイノルズ数の世界で生きており、ほとんど慣性力の影響を受けない。そのため、繊毛運動を停止すれば個体はすぐに停止することができる。しかし、細胞数が大きくなってきた多細胞緑藻は、慣性力の影響が無視できなくなる。
- 急に強い光を浴びることは光合成生物にとって大きなダメージとなる。そのため、その場に居続けることを避けるために緊急停止や後退遊泳をすることは、生存に重要な行動であると考えられる。ボルボックス目の藻類は、多細胞化(大型化)進化によって変化した流体環境でその行動を維持するために、同じ繊毛という運動器官の使い方を変化させていったと考えられる。
- 本研究の成果は、微生物の運動様式の進化を考える上で、流体環境の変化が淘汰圧になることを明確に示すものである。また、将来的には、ヒト繊毛病の発症メカニズムの理解を深めるとともに、バイオミメティック・マイクロマシンの設計に新たな展開をもたらすことが期待される。
研究体制
法政大学、京都産業大学の共同研究グループ
発表論文
「Multicellularity and increasing Reynolds number impact on the evolutionary shift in flash-induced ciliary response in Volvocales」
(多細胞化とレイノルズ数の増加がボルボックス目緑藻における閃光誘発繊毛運動調節の進化的シフトに与える影響)
背景
ボルボックス(図1の右2つ)は、教科書のカラー口絵にも掲載される、人気のある植物プランクトンの一種です。球状の個体がくるくる回りながら遊泳する様子は可愛らしく、多くの愛好家を惹きつけています。ボルボックスが属す緑藻植物門・緑藻綱・ボルボックス目(図1)にみられる多細胞性の生物群は、単細胞緑藻クラミドモナス(図1左端)に似た祖先単細胞が多細胞化することで進化してきたと考えられています。その証拠として、これらの生物は、どれも基本的には1細胞につき2本の繊毛(鞭毛)※1と呼ばれる運動器官と、1細胞につき1つの眼点※2と呼ばれる光受容器官を持つという、クラミドモナスが持つ特徴を共有しています。
ボルボックスの個体は、小さな種で数百、大きな種では数万もの細胞から成ります。一方、ボルボックス目の生物の中には、もっと少ない、さまざまな細胞数の種が含まれています。単細胞のクラミドモナスや、4細胞で1個体のテトラバエナ(シアワセモという和名でよく知られています)、16細胞が平板状に集まったゴニウム、16細胞が球状に集まったパンドリナ、32細胞が球状に集まったユードリナ…などの存在も、この生物群が共通単細胞祖先から多細胞化によって進化してきたことを示しています(図1)。生物の進化の歴史は、通常は化石を使って研究されます。しかし、ボルボックス目の生物は、2億年前という比較的「最近」多細胞化を始めたため、このように多様な細胞数の生物が現存しています。現存生物によって多細胞化進化の歴史を紐解くことができるというユニークな特徴から、「タイムマシン生物群」と呼ばれることもあります。
ボルボックス目の単細胞生物の代表であるクラミドモナスの一種、Chlamydomonas reinhardtii(以下、C. reinhardtii)は、光合成、繊毛運動、有性生殖など、広い分野でモデル生物とみなされています。C. reinhardtiiは、2本の繊毛をヒトの平泳ぎのように動かして水中を泳ぐ生物です。私達の研究グループは、これまで、C. reinhardtiiをはじめとするボルボックス目のいくつかの生物の光反応行動について研究してきました。C. reinhardtiiは、眼点で光を感受した刺激に応じて、平泳ぎをする際の2本の繊毛打の強さのバランスを変化させて走光性(光源に近づく正の走光性、光源から遠ざかる負の走光性)を示したり、平泳ぎからドルフィンキックのような繊毛波形に変換させて後退遊泳し、光驚動反応(急激な強い光刺激に対して運動を停止したり、遊泳方向を逆転させたりする行動)を示したりします(図2A)(参考文献1)。一方で、大型のボルボックスの一種 Volvox ferrisiiの場合は、走光性を示す際には前方の光源側の細胞の繊毛が、光驚動反応の際には前方のすべての細胞の繊毛が、それぞれ運動方向をほぼ逆転させます(図2B)(参考文献2)。さらに、4細胞のテトラバエナの一種 Tetrabaena socialisの場合、光を照射しても繊毛運動は顕著な変化を見せず、走光性も光驚動反応も示しません(参考文献3)。これらの現象から、私達は、ボルボックス目の生物の光反応行動における繊毛運動調節機構の多様性に興味を持ちました。

図1. ボルボックス目の藻類のいろいろ。ボルボックス目の多細胞化は、約2億年前という比較的「最近」始まり、徐々に細胞数を増やしていったと考えられている。これらの多様な細胞数の種は、段階的な多細胞化進化の歴史を反映するモデル生物群として注目されている。どの種も、1つの細胞に繊毛を2本と眼点を1つ持ち、水中を遊泳する。
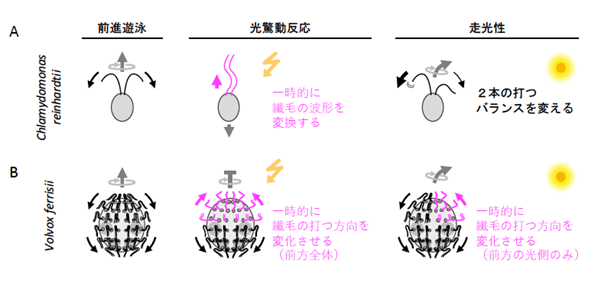
図2. A. 単細胞性のクラミドモナス(Chlamydomonas reinhardtii)と、B. 多細胞性のボルボックス(Volvox ferrisii)の光反応行動の際の繊毛運動様式変化。クラミドモナスもボルボックスも、通常は左のように、細胞または個体の後ろに向かって繊毛を打って前進する。急に周りの明るさが変わると、クラミドモナスは一時的な後退遊泳、ボルボックスは急停止する(光驚動反応)。一方向から光が当たり続けているときは、どちらも走光性を示す(この例では光源方向に向かう正の走光性)。クラミドモナスは2種類の繊毛の運動様式を使い分けることで2種類の光反応行動を行う。一方、ボルボックスはどちらの光反応行動でも繊毛運動様式は同じだが、反応する細胞の領域を変えることで2種類の光反応行動を行う。(黒またはマゼンタの矢印は繊毛の運動方向を、グレーの矢印は個体の運動方向を示す。)
研究成果
【ボルボックス目の27系統の生物の強光照射時の繊毛運動変化を観察】
そこで、ボルボックス目に属す、多様な細胞数の27系統が、光驚動反応を示すと期待される強光を照射したときの繊毛運動様式の変化を網羅的に解析しました。その結果、ボルボックス目藻類の強光照射時の繊毛運動様式は、4つのパターンに分類できることがわかりました(図3)。①波形変換、つまりクラミドモナスのように繊毛の打ち方を変えて一時的に後退遊泳するもの。②変化なし、つまりテトラバエナのように、強光を照射しても特に運動変化を見せないもの。③運動停止、つまり前方の細胞の繊毛運動が一時的に停止し、個体全体も停止するもの。④運動方向逆転、つまり前方の細胞の繊毛運動が、波形はそのままに、一時的に運動方向を逆転させ、個体全体も停止するもの。興味深いことに、この4つのパターンは、遺伝子解析からわかっている系統とは一致せず、ボルボックス目の中で互いに近縁な種が、必ずしも同じパターンを示すわけではありませんでした(図4)。
-
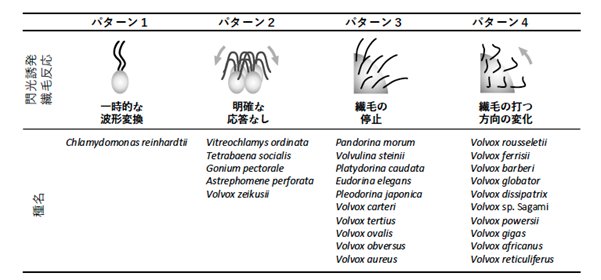
図3. ボルボックス目の26種27系統の強光照射時の繊毛運動変化は、4パターンに分類された。
-
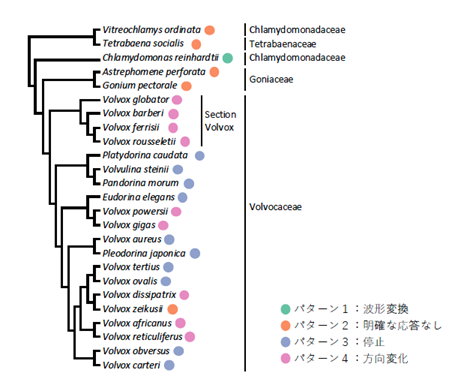
図4. 強光照射時の繊毛運動変化の4パターンは、系統関係とは完全には一致しなかった。
【レイノルズ数と繊毛運動パターンの連関】
しかし、別の指標から見てみると、このパターンに法則性が見えてきました。それが、生物が置かれている流体環境を示す「レイノルズ数」※3という概念です。レイノルズ数とは、運動している物体の慣性力※4を、その物体が受ける粘性抵抗※5で割った値です。たとえば、我々ヒトは、プールで壁を蹴った後、泳がなくてもしばらくの間は前に進むことができます(蹴伸び)。これは、ヒトが、水中において粘性抵抗よりも慣性力が大きい、つまりレイノルズ数が大きな環境にいることを示します。一方で、クラミドモナスは、レイノルズ数が微小な環境、すなわち、慣性力がほとんど無視できる環境に棲んでいます。クラミドモナスは、前進遊泳中に強光を感受した際、繊毛運動の波形変換を行うことで、急激に後退遊泳を行います(図2A)。クラミドモナスは前進遊泳の際、1秒間に約150 μm、体長の約15倍の距離を進みます(1 μm マイクロメートルは1 mmの1/1000の長さ)。これを自動車に置き換えてみると、時速約240 kmで走る車が、急にバックし始めるのと同じです。実際には、車のサイズではレイノルズ数が大きい、すなわち、慣性力が支配的なので、そのようなスピードで走る車は、バックはおろか、急に止まることすらできません。しかし、クラミドモナスのような微生物が棲む低レイノルズ数の世界では、粘性抵抗が支配的であり、慣性力がほとんどはたらきません。ヒトのサイズに置き換えると、クラミドモナスは、蜂蜜のようなネバネバの液体の中で泳いでいるようなものなのです。運動を止めればすぐ停止しますし、後ろ向きの力を発生させればすぐに後進します。
ボルボックス目の27系統の生物について、それぞれの生物の大きさと遊泳速度からレイノルズ数を算出すると、4つの繊毛運動変化パターンは、その系統ではなく、概ねレイノルズ数の変化に応じて変化していることがわかりました(図5)。すなわち、単細胞性の小さなクラミドモナスは、レイノルズ数が約0.001と、慣性力がほぼ無視できる流体環境におり、①繊毛波形変換によって後退遊泳することができます。多細胞化により少し大きくなったグループは、③前方の運動を停止させます。レイノルズ数は0.01〜0.1程度と、慣性力の影響が比較的小さい流体環境のため、後方に推進力が残っていても、個体は停止することができます。ところが、さらに大きくなり、レイノルズ数が0.1を上回るグループは、④前方の繊毛運動の方向を逆転させる、いわば「逆噴射」しています。これは、大きくなったことで徐々に慣性力が無視できない流体環境で生息することとなったため、強光照射に応じて急速に停止するために獲得した形質であると考えられます。また、②の特に変化のないグループのうち、以前私達が研究したテトラバエナは、強い光のストレスを感じない葉緑体機能を獲得する方向に進化したものでした。②に属す他の生物も、おそらく①、③、④とは別の生存戦略をとっていると考えられます。
同系統の生物は、基本的には大型化進化の道をたどると考えられています(「コープの法則」※6と呼ばれます)。大型化は、捕食者に食べられにくくなったり、生存に適した環境への移動が速くなったりなど、生存のための多くのメリットをもたらします。しかし、微細藻類の場合、多細胞化=大型化に伴い、周囲の流体環境が大きく変化するため、小さな祖先種が持っていた繊毛運動変化の形質をそのまま受け継ぐだけでは、光驚動反応のような重要な行動を示せなくなります。大型化に伴う流体環境変化(慣性力の増加)に適応するかたちで、ボルボックス目藻類は異なる繊毛運動変化パターンを獲得していったと考えられます。
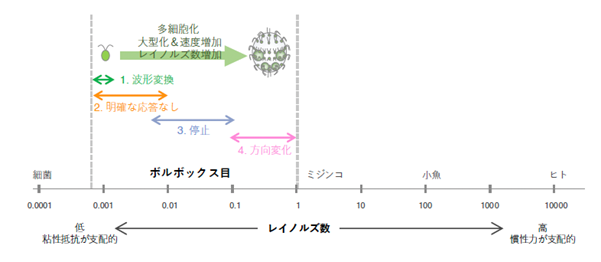
図5. レイノルズ数と繊毛運動パターンの連関。ボルボックス目に属す生物のレイノルズ数が多細胞化によって上昇するにつれて、繊毛運動パターンが1から4へと変遷することがわかった。(参考:S. Childress & R. Dudley 2004; K. Ishimoto 2012; 石本健太『微生物流体力学』2022)
今後の展望
本研究で、ボルボックス目の藻類が、多細胞化進化の過程で変化する流体環境にどのように適応してきたのか、その一端を明らかにすることができました。今後は、同じ繊毛という器官が、異なる運動様式変化を示す分子機構の解明が重要課題になります。繊毛は、微生物からヒトまでよく保存された細胞小器官であり、その運動調節が異常になることが原因のヒト疾患「原発性不動繊毛症候群」が知られています。繊毛運動が刺激に応じて運動様式を変化させることはヒト体内でも重要な役割を果たすため、ボルボックス目を用いて明らかになった運動変化パターンの調節分子機構がわかれば、原発性不動繊毛症候群の発症機構の理解に貢献すると期待されます。また、現在、広い分野で、小さなサイズの機械「マイクロマシン」「ナノマシン」の開発研究がされています。本研究の成果は、そうした小さな機械を操るのに必要な、サイズに応じた適切な運動調節システムを、バイオミメティクス(生物模倣)※7的に提案することにつながります。
責任著者コメント
近縁の微細藻類が、強光照射に応じて異なる繊毛運動調節パターンを示すことは、断片的には知られていましたし、また、これまでの私達の研究でも明らかにしてきました。今回の研究で、それを網羅的に解析することにより、レイノルズ数というキーワードと紐づけることができました。今後、近縁の生物が、保存性の高い繊毛という運動器官を、どのようにして異なる調節機構で操っているのか、分子レベルの知見を得ることに挑戦していきたいと考えています。
用語・事項の解説
※1 繊毛・鞭毛
真核細胞から生える、毛状の細胞小器官。動くものと動かないものがあるが、動かないものを一次繊毛と呼び、単純に繊毛・鞭毛と呼ぶ場合は動くものであることが多い。動物の精子の鞭毛のように、1細胞から1〜数本程度生えている場合は鞭毛、ゾウリムシのように、1細胞から多数生えている場合は繊毛と呼ぶが、相同の器官であるため、近年、繊毛と用語統一される動きがあり、本稿もそれに従う。
※2 眼点
多様な生物が持つ光受容器官だが、ボルボックス目緑藻の場合、細胞の赤道付近に1つ赤い点のように観察される。チャネルロドプシンと呼ばれる光駆動型イオンチャネルを細胞膜にもち、赤〜橙のカロテノイド色素がそれを裏打ちすることで、眼点の正面からの光のみを感受する、高指向性光受容機構をもつ。光を受容するとイオンを流入させ、細胞に光を受容したことを知らせる。
※3 レイノルズ数
流体力学の概念であり、ある球状の物体が、密度ρ(kg/m3)、粘性係数μ(kg/(m・s))の流体の中を、流れに対する相対平均速度V(m/s)でL運動しているとき、レイノルズ数ReはRe=ρV2/(μV/L)で与えられる。分子が慣性力を、分母が粘性抵抗を示す。
※4 慣性力
物体がそのままの状態(静止または等速直線運動)を続けようとする性質を慣性とよぶ。物体に力が加わって加速したり減速したりする時に、その物体がもとの状態を保とうとする力を慣性力とよぶ。(たとえば、車が急停止すると、乗っている人が元の速度で前に進もうとして、前に投げ出される時にはたらく力。)
※5 粘性抵抗
物体が液体や気体の中を動くとき、その物体が液体や気体から受ける抵抗力。(たとえば、水の中で手を動かすと、空気中で動かすよりも動きにくい。これは、水のほうが空気よりも粘性(ねばり)が強く、手が感じる粘性抵抗が高いため。)
※6 コープの法則
同じ系統の進化において、より大きなサイズの種がより新しい時代に出現する傾向があるという法則。提唱者Edward Drinker Copeの名に由来する。
※7 バイオミメティクス(生物模倣)
生物の構造や機能を模倣して、新しい技術や製品を開発する科学技術。(たとえば、蓮の葉の表面構造にヒントを得た撥水素材や、サメ肌の表面構造にヒントを得た水の抵抗の少ない水着など。)
論文情報
論文タイトル:Multicellularity and increasing Reynolds number impact on the evolutionary shift in flash-induced ciliary response in Volvocales(多細胞化とレイノルズ数の増加がボルボックス目緑藻における閃光誘発繊毛運動調節の進化的シフトに与える影響)
掲載誌 :国際学術誌BMC Ecology and Evolution(Impact Factor: 2.3)
掲載日時:2024年9月24日(日本時間)
著者:(1筆頭著者、2責任著者)1,2Noriko Ueki and 2Ken-ichi Wakabayashi
DOI:10.1186/s12862-024-02307-1
研究者一覧(所属先は2024年4月1日 現在)
植木紀子:法政大学自然科学センター・法学部法律学科教授
若林憲一:京都産業大学生命科学部産業生命科学科教授
謝辞
本研究は、日本学術振興会の科学研究費補助金(19K23758, 21K06295, 22H02642, 22H05674, 22H01440, 23K22711, 23K18136)、大隅基礎科学創成財団(2-G0008)、人・環境と物質をつなぐイノベーション創出ダイナミック・アライアンスの支援を受けて実施しました。
参考文献
1:10.1007/978-981-15-8763-4_2
2:10.1186/1741-7007-8-103.
3:10.1371/journal.pone.0259138
【研究に関するお問い合わせ先】
法政大学自然科学センター・法学部 教授 植木 紀子
TEL: 03-3264-4179 E-mail: noriko.ueki@hosei.ac.jp
【報道に関するお問い合わせ先】
法政大学 総長室広報課
TEL:03-3264-9240 E-mail:pr@adm.hosei.ac.jp